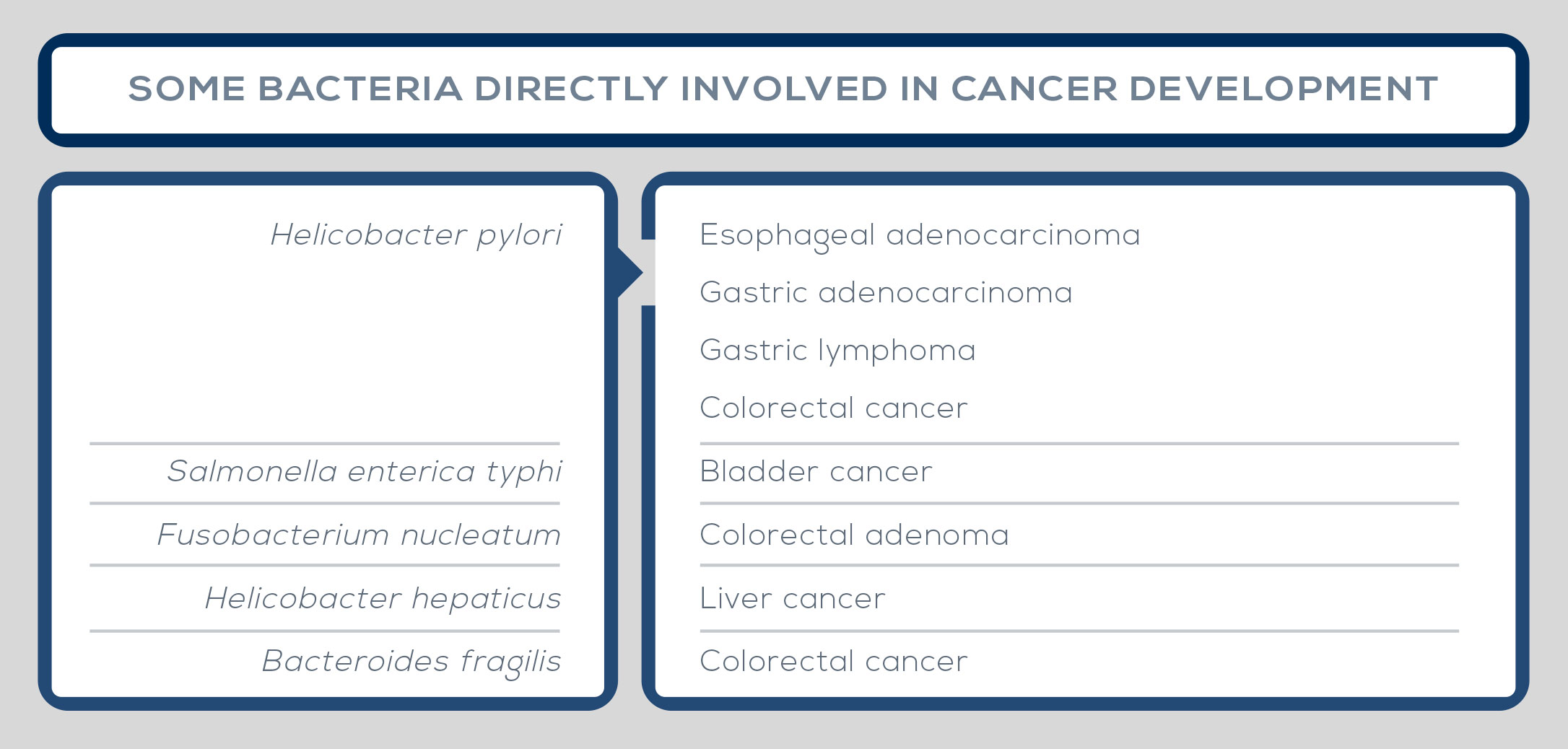
The microbiota exerts both local and systemic effects on cancer. In some cases, its composition promotes the development of the disease. However, some bacteria exert a protective effect, reducing the risk of cancer. Stomach, liver, and colorectal cancers stand as examples of the association between unfavorable microbiota features and cancer. In fact, stomach cancer development depends on the bacterium Helicobacter pylori, whereas the microbiota of colorectal cancer patients is characterized by increased Prevotella, and liver cancer has been linked to microbiota-derived bile acids that damage liver cell DNA.
But the list of cancers in which the microbiota has been implicated in the carcinogenic process is longer; among others are bladder carcinoma, lymphoma, prostate cancer, breast carcinoma, sarcoma, pancreatic cancer, and ovarian cancer. In some cases, single bacterial strains are responsible for driving the development of the disease, such as Helicobacter pylori in stomach cancer, Escherichia coli in colorectal carcinoma, and Salmonella enterica typhi in bladder carcinoma. Also, carcinogenesis can be locally modulated by groups of bacteria, which contribute to cancer development by interacting with the altered environment.
In other cases, cancer is driven by overall microbiota characteristics than by the presence of one species or one group of microbes, such as changes in bacterial density near mucous membranes. The vaginal microbiota has been linked to gynecological cancer, whereas organs with no known microbiota, such as liver, can be affected because of anatomical links with the gut allowing exposure to microbe-specific molecules (the so-called microorganismassociated molecular patterns, or MAMPs) and bacterial metabolites. Among mechanisms underlying the association between the microbiota and cancer development is the relationship between bacteria, inflammatory cytokines, and the immune system. Furthermore, some bacteria can affect cancer development by deregulating signals or pathways. For example, Helicobacter hepaticus promotes liver cancer by activation of NF-kB regulated networks.

Other bacteria directly affect carcinogenesis by producing virulence factors (such as toxins) that modulate inflammation in the cancer microenvironment, influence genomic stability of host cells, or epigenetically regulate host gene expression. For example, in colorectal cancer colibactin-synthesizing Escherichia coli is associated with DNA damage. Moreover, microbiota can interact with food and inflammation-related host metabolites that increase the growth of potential pathogens and the risk of DNA damage. It can, for example, enhance the production of secondary bile acids hypothesized to modify the mucosal barrier, allowing for antigen penetration and increasing inflammation and the production of factors needed for cancer development. Also, it can metabolize red meat and high processed food, giving rise to molecules believed to damage DNA, such as hydrogen sulfide. Finally, the correlation between obesity, a known risk factor for cancer, and gut microbiota is well documented, and aging, a well-known cancer driving factor, is associated with gut microbiota alteration as well.